


Antaŭaj esplorstudoj sugestas, ke L-aspartato, kiel L-glutamato, ekigas ekscitan agadon sur neŭronoj. L-aspartato funkcias kun L-glutamato en la sinaptaj vezikoj de nesimetriaj ekscitaj sinapsoj. Sed, la totala koncentriĝo de ĉi tiuj en la homa cerbo (0.96-1.62 ?mol/gramo malseka pezo), iliaj eksterĉelaj koncentriĝoj en la kortekso kiel mezurite per mikrodializo (1.62 ?M por L-aspartato kaj 9.06 ?M por L-glutamato) kaj ilia provizo laŭ imunohistokemio sugestas ke L-aspartato estas signife malpli abunda ol L-glutamato. Krome, L-aspartato estas potenca agonisto por NMDA-receptoroj sed ne por aliaj iGluR-oj kun EC50 nur ok-oble pli alta ol tiu de L-glutamato. EAAT-oj kiuj ludas fundamentan rolon en la asimilado de ĉiu vezikula liberigita L-glutamato en la centra nervosistemo (CNS) ankaŭ postulas la utiligon de L-aspartato. L-aspartato estas eble same malpli esenca kiel L-glutamato ligita al la totala ekscita agado asociita kun iGluRs. Kune kun ĝia rolo kiel neŭrotransmitoro, kiel antaŭe menciite, L-aspartato ankaŭ estas necesa kiel substrato por aspartata amino-transferazo kiu iĝas 2-oksoglutarato kaj L-glutamato por transporti al la kortikalaj vezikoj de glutamatergiaj neŭronoj kiuj ankaŭ povas sekve kaj nerekte pliigas liberigon de L-glutamato.
Enhavtabelo
Aliaj molekuloj en glutamato-signalado
Unu karakterizaĵo kiu distingas NMDA-receptorojn de malsamaj iGluRs estas ke la aktivigo de NMDA-receptoroj bezonas la ligon de ko-agonisto al la glicina ligregiono de la receptoro. Ekzemple, en la retino kaj en la mjelo, la origino de glicino povas disverŝi el glikinergiaj inhibiciaj sinapsoj. Sed, en malsamaj regionoj de la cerbo kun pliigita NMDA-receptor-esprimo, kiel ekzemple la hipokampa formado, mankas reagoj asociitaj kun striknin-sentemaj glicinaj riceviloj, almenaŭ en plenkreskaj neŭronoj, pruvante la foreston de glicinergic-inhibiciaj neŭrotransmisioj. Sed, glicino estas trovita en la eksterĉela likvaĵo de la hipokampo ĉe bazliniaj kvantoj de ĉirkaŭ 1.5 ?M, kiu estas simila al la saturiĝo de la glicina liganta regiono de la NMDA-receptoro, kvankam tiuj povas esti supren- kaj malsupren-reguligitaj. La origino de eksterĉela glicino en la hipokampo povas esti neŭronoj kiuj liberigas glicinon tra la alanin-serina-cisteina aminoacidtransportilo 1 (asc-1). Sed ankaŭ pruviĝis glicina liberigo de astrocitoj, kiu estas stimulita per malpolariĝo kaj kainato. Pliaj esplorstudoj estas postulataj por finfine montri ĉi tiujn rezultiniciatojn.
Eĉ en antaŭaj esplorstudoj de la NMDA-receptoro kaj ĝia kunaktivigo de glicino rivelis ke D-aminoacidoj, precipe D-serino, estas preskaŭ same potencaj kiel glicino. Nur kelkajn jarojn poste, evidentiĝis, ke D-serino troviĝas en rato kaj homa cerbo je proksimume triono de ilia koncentriĝo de L-serino havanta absolutan koncentriĝon de pli ol 0.2 ?mol/g cerba histo. Utiligante antiserumon por D-serino, esplorstudoj pruvis, ke D-serino el la cerbo troviĝas nur en astrocitoj kaj ĝia provizo konvenas al la esprimo de NMDA-receptoroj. Krome, la samaj esploristoj pruvis, ke D-serino estas liberigita de kulturitaj astrocitoj kiam eksponite al L-glutamato aŭ kainato. La abundo de D-serino estas trovita de la degradanta enzimo D-aminoacida oksidazo (DAO) kiu rivelas pliigitan esprimon en la postcerbo kie D-serina niveloj estas reduktitaj same kiel la sinteza enzimo serina racemase kiu kreas D-serinon de L- serino. D-Serino ŝajnas esti stokita en citoplasmaj vezikoj en astrocitoj kaj ĝi povas esti liberigita per ekzocitozo. Longperspektiva potencigo dependas de D-serina liberigo de astrocitoj en hipokampaj tranĉaĵoj, sugestante ke tiu aminoacido certe ludas fundamentan rolon en glutamatergia neŭrotranssendo tra NMDA-receptoroj. Aldone en hipokampaj tranĉaĵoj, esplorstudoj trovis, utiligante D-serinan kaj glicinan degradantajn enzimojn, kiuj D-serino funkcias kiel kundissendilo por sinaptaj NMDA-receptoroj sur CA1-neŭronoj same, kiuj glicino funkcias kiel la endogena ko-agonisto por ekstersinaptaj NMDA-receptoroj. Sinaptaj NMDA-receptoroj de dentitaj gironeŭronoj utiligas glicinon prefere ol D-serino kiel la ko-agoniston.
Prenitaj kolektive, multtavolaj rezultaj mezuroj montras, ke L-aspartato ne simple funkcias kiel agonisto sur NMDA-receptoroj sed ankaŭ glicino kaj D-serino ludas fundamentajn rolojn en glutamatergia neŭrotranssendo en la homa cerbo. Sed aliaj molekuloj ankaŭ pruviĝis esti signifaj moduliloj de glutamatergia neŭrotranssendo.
Glutamato Aktivigita de Aliaj Molekuloj
L-homocisteato (L-HCA) havas strukturajn similecojn kun L-glutamato. La ne-proteina aminoacido estas oksigenadprodukto de homocisteino kiu estas biosintezita de metionino en la elimino de sia propra fina metilgrupo kaj ĝi ankaŭ estas peranto de la transsulfuradpado de kiu metionino povas esti konvertita al cisteino tra cistationino. Fruaj esploraj studoj pruvis, ke ĉi tiu aminoacido povas kaŭzi kalcian enfluon en kulturitaj neŭronoj same sekure kaj efike kiel L-glutamato. Plie, L-HCA rivelis pliigitan afinecon por NMDA-receptoroj kompare kun aliaj iGluR-oj en devigaj provoj asociitaj kun ĝia kapablo kaŭzi NMDA-receptoron-antagonist-inhibiteblan ekscitotoksecon kaj natrienfluon. Aldone, L-HCA povas ekigi mGluR5 same efike kiel L-glutamato. L-HCA troviĝas en la cerbo, tamen, la koncentriĝoj pruviĝis proksimume 500-oble pli malgrandaj ol tiuj de L-glutamato kaj eĉ 100-oble pli malgrandaj kompare kun tiuj de L-aspartato en malsamaj regionoj de la ratcerbo. Dum kalio-induktita stimulo, L-HCA-senŝargiĝo estas ekigita de cerbaj tranĉaĵpreparoj kiel montrite por L-aspartato kaj L-glutamato kvankam la absoluta liberigo de HCA estas proksimume 50-oble pli malgranda. Surprize, HCA estas tre efika konkurenciva inhibitoro de cistino kaj L-glutamato-asimilado per la cistina/glutamato kontraŭportera sistemo x?c, la agado kiu reguligas kaj administras la eksterĉelajn ekstrasinaptajn L-glutamato-koncentriĝojn en la cerbo. Tial, la efiko de L-HCA sur la aktivigo de NMDA kaj aliaj L-glutamato-receptoroj ankaŭ povas dependi de la L-HCA-induktita ellasilo de L-glutamato per sistemo x?c. L-HCA povas ludi gravan rolon en la ĝenerala stimulo de L-glutamato-receptoroj. Tamen, tio povas ege ŝanĝiĝi sub certaj kondiĉoj, ekz., en pacientoj kun alt-doza metotreksatterapio, kontraŭkancera medikamento kiu, limigante dihidrofolatreduktazon, limigas la tetrahidrofolat-katalizitan recikladon de metionino de homocisteino. Ĉi tie, L-HCA-koncentriĝoj de pli ol 100 ?M estis pruvitaj de la cerbo-spina likvaĵo dum L-HCA estis nerimarkebla en kontrolsubjektoj. Pliaj esplorstudoj daŭre estas postulataj por determini ĉi tiujn rezultiniciatojn.
Pliaj endogenaj malgrandaj molekuloj, kiuj supozeble influas signaladon de L-glutamato, inkluzivas plurajn intermediarojn de triptofanmetabolo, kiel montrite en Figuro 2. Tra la agado de indoleamino 2,3-dioksigenazo (IDO) aŭ triptofano 2,3-dioksigenazo (TDO), triptofano estas igita N-formil-L-kinurenino kiu poste estas igita kinurenino (KYN) per formamidazo. Tri vojoj, du el kiuj ligas ĉe posta paŝo, rezultigas plian metabolon. Unue, per la agado de kinurenina aminotransferazo (KAT), KYN estas konvertita en kinurenan acidon (KYNA). KYN ankaŭ povas esti konvertita al 3-hidroksikinurenino (3HK) per kinurenina monooksigenazo (KMO), kiu poste povas esti utiligita kiel substrato per kinureninazo por la sintezo de 3-hidroksiantranila acido (3HANA). Plie, utiligante KYN kiel substraton, kinureninazo evoluigas antranilacidon (ANA), kiu per nespecifa hidroksilation ankaŭ povas esti konvertita al 3HANA. Laŭ esplorstudoj, 3HANA finfine funkcias kiel substrato por la generacio de kinolina acido (QUIN).
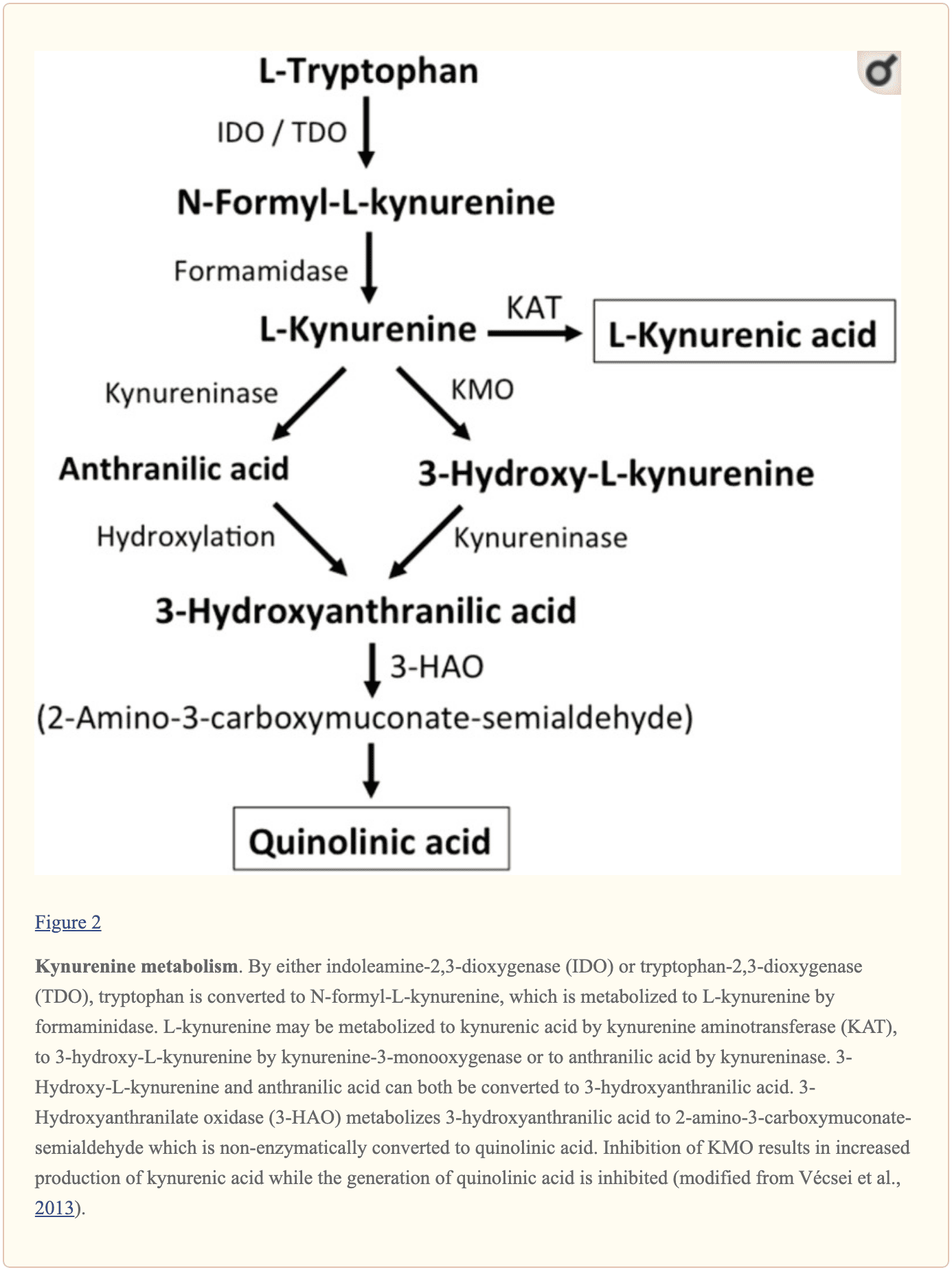
La triptofankoncentriĝo en la ratcerbo estas ĉirkaŭ 25 nmol/g malseka pezo kaj proksimume 400-oble malpli ol L-glutamato kaj 100-oble malpli ol L-aspartato. La pruvitaj cerbaj niveloj de kinureninoj estas eĉ pli malaltaj kun 0.4-1.6 nmol/g por QUIN, 0.01-0.07 nmol/ml por KYNA, kaj 0.016 nmol/g por 3HANA. Ĉirkaŭ 40 procentoj de cerbo KYN estas loke sintezitaj. La metabolitoj de triptofano montras diferencigan ligon al plasmoproteinoj kaj sian transporton tra la bariero kiu estas tre malsama. KYN kaj 3HK estas portitaj tra la granda neŭtrala aminoacida portanta sistemo L. Kynurenines ŝajnas penetri la homan cerbon per pasiva difuzo. Plie, KYNA, 3HANA, kaj precipe ANA ligas al serumproteinoj kiuj tiam finfine limigas kaj limigas sian difuzeblecon trans la sango-cerba baro.
Esplorstudoj pruvis ke QUIN, kiam jonoforetike utiligita en ratĉeloj, kaŭzis neŭronan pafadon kiu estis malhelpita fare de NMDA-receptoantagonisto, sugestante ke QUIN povas funkcii kiel NMDA-recepto-agonisto. Tamen, la EC50 por QUIN por ekigi NMDA-ricevfluojn montriĝis proksimume 1000-oble pli alta ol la EC50 de L-glutamato. Encerba injekto de QUIN estis pruvita kaŭzi ultrastrukturajn, neŭrokemiajn, kaj kondutismajn ŝanĝojn similajn al tiuj kaŭzitaj de NMDA-receptoraj agonistoj. La fakto ke QUIN-koncentriĝoj estas proksimume 5000- ĝis 15,000-oble pli malaltaj ol cerbaj L-glutamato-koncentriĝoj faras malprobabla ke modulado de NMDA-receptorsignalado de QUIN ludas esencan rolon. KYNA pruviĝis funkcii kiel NMDA receptorantagonisto. Sed, kvankam infuzaĵo kun la KMO-inhibitoro Ro 61-8048 plibonigis cerbajn eksterĉelajn KYNA-koncentriĝojn 10-oble, tio ne rezultigis inhibicion de NMDA-mediaciita neŭrona malpolariĝo, trovo kiu defias la kredon ke KYNA ĉe preskaŭ fiziologiaj kvantoj rekte. modulas NMDA-receptorojn. Kompare, pliigita KYNA en la cerbo induktita de la KMO-inhibitoro JM6 malpliigis la eksterĉelan cerban L-glutamato-koncentriĝon. Plie, KYNA-niveloj de la eksterĉela cerba likvaĵo estis asociitaj kun L-glutamatniveloj sugestante ke eĉ sur fiziologiaj aŭ proksime de fiziologiaj niveloj, KYNA modulas L-glutamatmetabolon. Kaj la aktivigo de la G-protein-kunligita receptoro GPR35 kaj la inhibicio de presinaptaj ?7 nikotinaj acetilkolinreceptoroj estas sugestitaj en la KYNA-induktita redukto en L-glutamato liberigo. Por resumi, kvankam QUIN kaj L-HCA ĉeestas en la homa cerbo, iliaj koncentriĝoj diskutas kontraŭ ili kun roloj en reguligado kaj konservado de neŭrotranssendo. En kontrasto, eĉ se la vojoj devas esti difinitaj pli detale, indico subtenas nivelojn kaj la opinion ke senŝargiĝo povas esti modulita per KYNA kaj neŭrotranssendo.

Glutamato, kune kun aspartato kaj aliaj molekuloj, estas pluraj el la ĉefaj ekscitaj neŭrotransmisiloj en la homa cerbo. Kvankam ĉi tiuj ludas fundamentan rolon en la ĝenerala strukturo kaj funkcio de la centra nervosistemo, inkluzive de la cerbo kaj la mjelo, troaj kvantoj de aliaj molekuloj povas finfine ekigi glutamatajn ricevilojn. Troa glutamato povas kaŭzi ekscitotoksecon, kiu povas konduki al diversaj sanproblemoj, kiel Alzheimer-malsano kaj aliaj specoj de neŭrologiaj malsanoj. La sekva artikolo priskribas kiel aliaj molekuloj povas aktivigi glutamatajn receptorojn. – D-ro Alex Jimenez DC, CCST Insight – D-ro Alex Jimenez DC, CCST Insight
Neuropatia Traktado kun LLLT
Esploraj studoj sugestas, ke L-aspartato, kiel L-glutamato, ekigas ekscitan agadon. L-aspartato funkcias kun L-glutamato en la sinaptaj vezikoj de nesimetriaj ekscitaj sinapsoj. Sed, la totala koncentriĝo de ĉi tiuj en la homa cerbo sugestas, ke L-aspartato estas signife malpli abunda ol L-glutamato. Krome, L-aspartato estas potenca agonisto por NMDA-receptoroj sed ne por aliaj iGluR-oj kun EC50 nur ok-oble pli alta ol tiu de L-glutamato. La amplekso de niaj informoj estas limigita al kiropractikaj, muskoloskeletaj kaj nervozaj sanaj aferoj kaj ankaŭ al funkciaj medicinaj artikoloj, temoj kaj diskutoj. Ni uzas funkciajn sanprotokolojn por trakti vundojn aŭ kronikajn malordojn de la muskoloskeleta sistemo. Por plu diskuti la ĉi-supran temon, bonvolu demandi doktoron Alex Jimenez aŭ kontakti nin ĉe 915-850-0900 .
Kuraĝita de doktoro Alex Jimenez
Referencoj
- Lewerenz, Jan, kaj Pamela Maher. "Kronika Glutamata Tokseco en Neŭrodegeneraj Malsanoj - Kio Estas la Indico?" Frontejoj en Neŭrikeco, Frontiers Media SA, 16 Dec. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4679930/.
Pliaj Temaj Diskutoj: Kronika Doloro
Subita doloro estas natura respondo de la nerva sistemo, kiu helpas pruvi eblajn vundojn. Ekzemple, doloraj signaloj veturas de vundita regiono tra la nervoj kaj spina ŝnuro ĝis la cerbo. Doloro estas ĝenerale malpli severa dum la vundo resanigas, tamen kronika doloro estas malsama ol la averaĝa speco de doloro. Kun kronika doloro, la homa korpo daŭre sendos dolorajn signalojn al la cerbo, sendepende de se la vundo resaniĝis. Kronika doloro povas daŭri de pluraj semajnoj ĝis eĉ pluraj jaroj. Kronika doloro povas ege influi la moveblecon de paciento kaj ĝi povas redukti flekseblecon, forton kaj persistemon.
Neural Zoomer Plus por Neŭrologia Malsano

D-ro Alex Jimenez uzas serion de provoj por helpi taksi neŭrologiajn malsanojn. La Neŭra ZoomerTM Plus estas tabelo de neŭrologiaj aŭtorokorpoj, kiuj ofertas specifan rekonon de antikorpoj al antigenoj. La Vibra Neŭra ZoomerTM Plus estas desegnita por taksi la reakcion de individuo al neŭrologiaj antigenoj de 48 kun ligoj al vario de neŭrologie rilataj malsanoj. La Vibra Neŭra ZoomerTM Plus celas redukti neŭrologiajn kondiĉojn ebligante pacientojn kaj kuracistojn kun esenca rimedo por frua riska detekto kaj plibonigita fokuso pri personigita primara antaŭzorgo.
Formuloj por Metilada Subteno

XYMOGEN Ekskluzivaj Profesiaj Formuloj estas haveblaj per elektitaj rajtigitaj sanitaraj profesiuloj. La interreta vendo kaj rabatado de XYMOGEN-formuloj estas strikte malpermesitaj.
Fiera, D-ro. Aleksandro Jimenez faras XYMOGEN-formulojn haveblajn nur al pacientoj sub nia zorgo.
Bonvolu nomi nian oficon por ke ni asignu kuraciston konsulti por tuja aliro.
Se vi estas pacienca Medicina kaj Kiropractika Kliniko de Vundo, vi povas demandi pri XYMOGEN vokante 915-850-0900.
![]()
Por via komforto kaj revizio pri la XYMOGENO produktoj bonvolu revizii la sekvan ligon. *XYMOGEN-Katalogo-Elŝuti
* Ĉiuj supre menciitaj politikoj XYMOGEN restas strikte validaj.
Afiŝu Malgarantion
La informoj ĉi tie en "Funkcia Neŭrologio: Aliaj Molekuloj en Glutamato" ne celas anstataŭigi unu-kontraŭ-unuan rilaton kun kvalifikita sanprofesiulo aŭ licencita kuracisto kaj ne estas medicina konsilo. Ni instigas vin fari sandecidojn bazitajn sur via esplorado kaj partnereco kun kvalifikita sanprofesiulo.
Blogaj Informoj kaj Ampleksaj Diskutoj
Nia informa amplekso estas limigita al Kiropractiko, muskoloskeleta, akupunkturo, fizikaj medikamentoj, bonfarto, kontribuanta etiologia viscerosomataj perturboj ene de klinikaj prezentoj, asociita somatoviscera refleksa klinika dinamiko, subluksaj kompleksoj, sentemaj sanproblemoj, kaj/aŭ funkciaj medicinaj artikoloj, temoj kaj diskutoj.
Ni provizas kaj prezentas klinika kunlaboro kun specialistoj el diversaj fakoj. Ĉiu specialisto estas regita de ilia profesia amplekso de praktiko kaj ilia jurisdikcio de licenco. Ni uzas funkciajn sanajn kaj bonfartajn protokolojn por trakti kaj subteni prizorgon por la vundoj aŭ malordoj de la muskoloskeleta sistemo.
Niaj filmetoj, afiŝoj, temoj, temoj kaj komprenoj kovras klinikajn aferojn, aferojn kaj temojn, kiuj rilatas al kaj rekte aŭ nerekte subtenas nian klinikan amplekson de praktiko.*
Nia oficejo prudente provis provizi subtenajn citaĵojn kaj identigis la koncernajn esplorstudojn subtenantajn niajn afiŝojn. Ni provizas kopiojn de subtenaj esploraj studoj haveblaj al reguligaj estraroj kaj publiko laŭ peto.
Ni komprenas, ke ni traktas aferojn, kiuj postulas plian klarigon pri tio, kiel ĝi povas helpi en aparta prizorgoplano aŭ traktado-protokolo; tial, por plu diskuti la temon supre, bonvolu bonvolu demandi D-ro Alex Jimenez, DC, Aŭ kontaktu nin ĉe 915-850-0900.
Ni estas ĉi tie por helpi vin kaj vian familion.
benoj
D-ro. Alex Jimenez A.D, MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
retpoŝto: trejnisto@elpasofunctionalmedicine.com
Licencite kiel Doktoro pri Kiropraktiko (DC) en Teksaso & Nov-Meksiko*
Teksasa DC-Licenco # TX5807, Nov-Meksiko DC Licenco # NM-DC2182
Licencite kiel Registrita Flegistino (RN*) en Florido
Florida License RN License # RN9617241 (Kontrolo Nr. 3558029)
Kompakta Statuso: Plurŝtata Licenco: Rajtigita Praktiki en 40-ŝtatoj*
Nuntempe Matrikulita: ICHS: MSN* FNP (Programo pri Familio Flegistinisto)
D-ro Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Mia Cifereca Komerckarto
 Denove Ni Bonvenigas Vin¸
Denove Ni Bonvenigas Vin¸
